J. Ge, Y. Hu, C. Ren, L. Guo, C. Wang, W. Sun, M.H. Shahrajabian
Abstract. Oat seed with dormancy characteristics, which can germinate after one season or one year, are used to build and maintain vegetation to protect soils from been damaged by desertification in Northern China. The aim of this study was to estimate the effects of endogenous and exogenous GA3 and ABA on oat seed (var. Baiyan 7) germination. The results showed that seeds without peel hull had lower endogenous ABA content and the ratio of ABA/GA3 than seeds with peel hull. The best GA3 treatment duration for milky ripe, wax ripe, full ripe seeds were 60 min or 120 min, 60 min and 30 min, respectively. Seed germination rate, germination potential and germination index increased before they declined with the increasing of GA3 concentrations. The best GA3 concentration treatment was 100 mg/l, while the turning point was 200 mg/l. The dormancy rate of low temperature storage seeds were higher than those of room temperature storage seeds at each storage time, and both decreased with the increase of the storage time. For the seeds which were new or stored for 1-2 months, the germination rates were enhanced significantly by exogenous GA3. For the seeds that had been stored for over three months, GA3 treatment had no effect on germination rate. Germination rate decreased with the increase of ABA concentrations. The most inhibitive effect, which leaded to a seed germination reduction by 37.7% and 4.0%, appeared, when the concentration of ABA was 500 mg/L and 1000 mg/l, respectively. GA3 could abate the effect which ABA inhibited seed germination.
Keywords: dormancy oats; Baiyan; germination index; germination potential.
View full article (HTML)
Effects of GA3 and ABA on the germination of dormant oat seeds
J. Ge1, Y. Hu2, C. Ren3, L. Guo3, C. Wang3, W. Sun4,5, M.H. Shahrajabian2,4,5*
1Zhangjiakou Academy of Agricultural Sciences, Zhangjiakou City, Hebei, China
2College of Agronomy and Biotechnology, China Agricultural University, Beijing, China
3Baicheng Academy of Agricultural Sciences, Baicheng City, Jilin, China
4Biotechonoly Research Institute, Chinese Academy of Agricultural Science, Beijing, China
5Nitrogen Fixation Laboratory, Qi Institute, Chuangye Country, Jiaxing, Zhejiang, China
*E-mail: hesamshahrajabaian@gmail.com
Received Feb. 27, 2018. Revised: May 14, 2018. Accepted: June 28, 2018. Published online: July 17, 2019
Abstract. Oat seed with dormancy characteristics, which can germinate after one season or one year, are used to build and maintain vegetation to protect soils from been damaged by desertification in Northern China. The aim of this study was to estimate the effects of endogenous and exogenous GA3 and ABA on oat seed (var. Baiyan 7) germination. The results showed that seeds without peel hull had lower endogenous ABA content and the ratio of ABA/GA3 than seeds with peel hull. The best GA3 treatment duration for milky ripe, wax ripe, full ripe seeds were 60 min or 120 min, 60 min and 30 min, respectively. Seed germination rate, germination potential and germination index increased before they declined with the increasing of GA3 concentrations. The best GA3 concentration treatment was 100 mg/l, while the turning point was 200 mg/l. The dormancy rate of low temperature storage seeds were higher than those of room temperature storage seeds at each storage time, and both decreased with the increase of the storage time. For the seeds which were new or stored for 1-2 months, the germination rates were enhanced significantly by exogenous GA3. For the seeds that had been stored for over three months, GA3 treatment had no effect on germination rate. Germination rate decreased with the increase of ABA concentrations. The most inhibitive effect, which leaded to a seed germination reduction by 37.7% and 4.0%, appeared, when the concentration of ABA was 500 mg/L and 1000 mg/l, respectively. GA3 could abate the effect which ABA inhibited seed germination.
Keywords: dormancy oats; Baiyan; germination index; germination potential.
INTRODUCTION
China’s desertification area is about 1.74 million km2, accounting for 18% of the total land area, and the area is expected to increase in the future (Chang et al., 2005; Islam et al., 2011; Chen et al., 2013). To solve this problem, it is important to build and maintain plant biology to increase surface coverage and restore native vegetation, which is the main reason why Chinese government has set up China Agriculture Research System to support the production and study of oat (Lin et al., 2012). Oat (Avena sativa L.) is gramineous precocious subfamily of annual grain oats herbaceous plants. It has the characteristics of strong ability to resist wind erosion and easy to sow, and easy to build plant (Zhao and Tian, 2007; Peiretti et al., 2011; Lin et al., 2012; Soleymani and Shahrajabian, 2012; Soleymani et al., 2012; Yong et al., 2017); suitable planting areas are similar with the current climatic conditions of the desertification area (Ren et al., 2007; Rabiei et al., 2012; Ogbaji et al., 2013; Shahrajabian et al., 2017).
Li et al. (2009) showed that the ability of anti-erosion oat is stronger than corn, sunflower and mung bean, which will help prevent farmland desertification. “Baiyan 7”, breed by Baicheng Academy of Agricultural Sciences, has dormancy characte-ristics. Some other researches showed the dormancy strength of “Baiyan 7” can affect the vegetation establish-ment.
Now, the studies of oat planting have focused on breeding, cultivation, physiological and biochemical aspects; oat seed science research focuses on the germination optimum temperature, germination substrate, salinity stress and seed storage, but the researches about the effect of phytohormones on germination of dormant oat arequite few. So, the study on plant hormones effect on oat dormancy has great practical importance for guide practice operations and seed breeding, provide the basis of the germination ecology in arid areas about vegetation recovery and reconstruction (Facteau et al., 1992). Khan (1975) put forward three factor hypothesis about seed germination hormone, GA3 is the main regulator, and practice has also been shown, GA3 could lift the bud and seed dormancy and promote bud, can instead of light or low temperature some seed break dormancy needs (Chen and Huang, 1998; Mukhtar and Singh, 2006; Ozkaya et al., 2006). Beck and Ziegler (1989) believed that germinated brown rice in addition to the need of water and a certain temperature, GA3 was a promoting substance of seed germination. ABA is a phytohormon, which has been shown to be involved in a wide range of plant physiology (Wilkinson and Davies, 2010). ABA inhibition of seed germination requirement concentration varies. Thus, the aim of this study is to estimate the effects of GA3 and ABA on germination of oat seeds.
MATERIAL and METHODS
Study site and materials
The experiment took place at Baicheng Academy of Agricultural Sciences, which is located in Baicheng, Jilin province, China (45° 37ˊN, 122° 48ˊE, 152 m elevation) in 2015. The oat seed used is “Baiyan 7”, collected on September 10th, bred by the Baicheng Academy of Agricultural Sciences. Part of the seeds was collected immediately for germination test, and other stored at room temperature under dry conditions or at 4℃ refrigerator for later use.
Experimental design
a) The effects of GA3 on germi-nation of oat seed in different maturity
Each different maturity oat seed were soaked with 100mg/l GA3 in different times to test germination. The treatments time were 0 min, 30 min, 60 min, 120 min, 240 min; different maturity were milky ripe, wax ripe, and full ripe. This experiment had 15 treatments; each treatment was repeated three times with 200 seeds in each repeat.
First, the seeds soaked and disin-fected in 4.0% sodium hypochlorite solu-tion for 30 min, then washed 5-6 times with distilled water.
Second, tweezers were used to put the seeds in Petri dishes lined with filter paper, each plate contained 200 seeds and were kept at sufficient distance.
Third, the Petri dishes were placed into 20℃ incubator, distilled water was used to keep the filter moist, every day sprout numbers were recorded on each treatment, for 10 days, each process repeated three times (These materials and methods are applicable to the following experiments).
b) The effects of GA3 concen-trations on wax ripe seed germination
The wax ripe seeds were soaked with different GA3 concentrations for
120 min, and test the germination. This experiment had six treatments: CK0 mg/l, T112.5 mg/l, T225 mg/l, T350 mg/l, T4100 mg/l and T5200 mg/l.
c) The effects of GA3 on germi-nation of oat seed in different maturity, with or without peel
Different maturities were cream ripe, wax ripe and full ripe. Each maturity seeds had four treatments: CK the seeds with peel; T1 the seeds without peel; T2 the seeds with peel, soaked with GA3 for 2 h; T3 the seeds without peel, soaked with GA3 for 2 h.
d) The effects of GA3 on germi-nation of oat seed in different maturity with low or room temperature
The seeds with different maturities were stored in low or room temperature. Different maturities were cream ripe, wax ripe and full ripe. Each kind of seeds had five storage durations: 0 month, 1 month, 2 month, 3 month and 4 month.
e) The interaction effect of GA3 and ABA on germination of oat seed
GA3 and ABA were dissolved in a small amount of ethanol, constant volume with distilled water, GA3 preparation of 100 mg/l, the ABA 1000 mg/l as stock solution, respectively, using the GA3 liquid and different concentrations of ABA dilution to deal with the sterilized seed, soak 30 min, remove seeds, and dry with filter paper, than do germination test. If GA3 and ABA are used, first the seed soak with different concentrations of ABA solution 30 min, after that soak with GA3 solution for 30 min, and then dry with filter paper and do germination test. This experiment has nine treatments: 1000 mg/l ABA (T1); 500 mg/l ABA (T2); 250 mg/l ABA (T3); 100 mg/l ABA (T4); 100 mg/l GA3+100 mg/l ABA (T5); 100 mg/l GA3+250 mg/l ABA (T6); 100 mg/l GA3+500 mg/l ABA (T7); 100 mg/l GA3+1000 mg/l ABA (T8); CK use distilled water as control.
Determination indexes and methods
a) Seed collection standard
Oat seed development process was divided into three maturities. Full ripe: glumes white and open, the appearance of seed is yellowish-white, hard; wax ripe: glumes the sallow semi-open, the appearance of seed is yellow-green, slightly harder, volume reach mature state; milk ripe: glumes green and closed, the appearance of seed is green, tender, the volume does not reach mature state.
b) Determination of germination rate, germination potential, germinating, germination index and T50
Test method reference to the international seed testing and GB/T2930.4-2001.
Germinating refers to the ratio of the sum the maximum number of germination of three days and total number of germination.
T50 refers to seed germination rate of the time required in half of the final germination.
Germination rate (%) = (n/N) ×100 (1)
In this equation, n refers to the seed within the specified time normal germination accumulated grains and N refers to the total number of tested seeds.
Germination potential (%) = (A/N) ×100 (2)
In equation number 2, A refers to the cumulative germination rate of 3d before the test.
Germination index = ΣGt/Dt, (3)
In equation number 3, Gt refers to the number of germination of the time t and Dt refers to the germination days.
Analyses
Experimental raw data use the Excel (2010 version) statistical software to collate, then use both SAS (8.0 version) and Mstatc-C statistical software to analyse.
A factorial layout within rando-mized complete block design with three replications was used for analysis variation the effect of GA3 processing time on germination characteristics of seeds in different maturity.
Seed maturity were milky ripe seed, wax ripe seed and full ripe seed. GA3 processing times were included 0, 30, 60, 120 and 240 min. In order to determine the influence on GA3 concentration on different experimental characteristics, a randomized complete block design with three replications were used. GA3 concentrations were 0, 12.5, 25, 50, 100 and 200 mg/l.
RESULTS AND DISCUSSION
As shown in Table 1, full ripe seeds, with or without peel, had significant differences (p<0.05) in endogenous GA3, ABA and GA3/ABA, at different storage periods.
Endogenous GA3 in full ripe seeds without peel in each storage period were significantly higher than those with peel (p<0.05).
With the extension of storage time, endogenous GA3 in full ripe seeds without peel were increased by 68.7%, 121.4%, 59.2%, 18.2%, and 29.0%, respectively, when compared with seeds with peel. GA3 contents showed a big increase trend with the storage time.
Endogenous ABA in full ripe seeds with peel in each storage period was significantly higher than that in the seeds without peel (p<0.05), with the extension of storage time; endogenous ABA in full ripe seed with peel than seed without peel were increased by 8.4%, 49.0%, 37.7%, 74.5%, and 36.7%, respectively, which means that it contains a certain amount of ABA within the peel.
The ratio of GA3/ABA in full ripe seeds without peel in each storage period was significantly higher than that in the seeds with peel (p<0.05), with the extension of storage time, the ratio of GA3/ABA in full ripe seeds without peel than seed with peel were increased by 82.8%, 229.9%, 119.2%, 106.2%, and 76.3%, respectively (Table 1).
Table 1
Full ripe seed phytohormones content at different storage periods with or without peel
|
Treatment |
Determination index |
Storage time (month) |
||||
|
0 |
1 |
2 |
3 |
4 |
||
|
With peel |
GA3 (ng/g.FW) |
9.25db |
5.24eb |
11.50cb |
19.18bb |
39.95ab |
|
Without peel |
15.61da |
11.60ea |
18.31ca |
22.67ba |
52.52aa |
|
|
With peel |
ABA (ng/g.FW) |
146.18ca |
137.41ea |
148.42ba |
169.33aa |
138.01da |
|
Without peel |
134.91ab |
92.23eb |
107.81bb |
97.03db |
100.94cb |
|
|
With peel |
GA3/ABA |
0.06db |
0.04eb |
0.08cb |
0.11bb |
0.29ab |
|
Without peel |
0.12da |
0.13da |
0.17ca |
0.23ba |
0.51aa |
|
Different letters within a row indicate significant differences between the mean (p<0.05)
The results showed that wax ripe seed, with or without peel, have significant differences (p<0.05) in endogenous GA3, ABA and GA3/ABA, at different storage periods.
Wax ripe seed without peel, GA3 content in addition to storage 3 months was significant less than seed with peel (p<0.05); the rest are significantly higher than wax ripe seed with peel (p<0.05), with the storage time of 0 min (month), 1 min, 2 min, 4 min, and endogenous GA3 in wax ripe seed without peel than seed with peel, the increase by 67.7%, 15.5%, 80.1%, and 59.9% was seen. Endogenous ABA in wax ripe seed with peel in each storage period are significantly higher than that without peel (p<0.05), with the extension of storage time; endogenous ABA in wax ripe seed with peel than seed without peel were increased by 55.6%, 152.8%, 165.9%, 270.9%, and 72.9%, respectively; this means that it contains a certain amount of ABA within the peel.
The ratio of GA3/ABA in wax ripe seed without peel in each storage period are significantly higher than seed with peel (p<0.05).
Moreover, with the extension of storage time, the ratio of GA3/ABA in full ripe seed without peel than seed with peel were increased by 161.0%, 191.9%, 378.8%, 209.9%, 176.3% (Table 2).
Table 2
Wax ripe seed phytohormones content of the different storage period with or without peel
|
Treatment |
Determination index |
Storage time (month) |
||||
|
0 |
1 |
2 |
3 |
4 |
||
|
With peel |
GA3 (ng/g.FW) |
19.79cb |
20.10cb |
16.45db |
23.12ba |
37.09ab |
|
Without peel |
33.20ba |
23.20da |
29.63ca |
19.32eb |
59.29aa |
|
|
With peel |
ABA(ng/g.FW) |
223.03aa |
158.33da |
169.16ca |
202.80ba |
131.84ea |
|
Without peel |
143.37ab |
62.63db |
63.62cb |
54.67eb |
76.27bb |
|
|
With peel |
GA3/ABA |
0.09db |
0.13bb |
0.10cdb |
0.11bcb |
0.28ab |
|
Without peel |
0.23da |
0.37ca |
0.47ba |
0.35c |
0.78aa |
|
Different letters within a row indicate significant differences between the mean (p<0.05).
The results showed that milky ripe seed with peel or without peel, the endogenous GA3, ABA and GA3/ABA, at different storage periods, have significant differences (p<0.05). Milky ripe seed without peel GA3 content in addition to storage for 4 months had no significant with seed with peel (p<0.05), the rest were significantly higher than seed with peel (p<0.05), with the storage time of 0 m (month), 1 min, 2 min, 3 min, endogenous GA3 in milky ripe seed without peel than seed with peel were increased by 44.0%, 37.7%, 76.9%, and 25.7%, respectively; the endogenous ABA in milky ripe seed with peel in each storage periods are significantly higher than seed without peel (p<0.05), with the extension of storage time; endogenous ABA in milky ripe seed with peel than seed without peel were increased by 38.9%, 18.8, 162.2, 107.9, 71.3%; this means it contain a certain amount of ABA within the peel. Milky ripe seed without peel the ratio of GA3/ABA in addition to storage for 0 month had no significant difference with the seed with peel (p<0.05), the rest were significant (p<0.05); with the storage time of 2 min (month), 3 min, 4 min, and the ratio of GA3/ABA in milky ripe seed without peel than seed with peel were increased by 48.2%, 65.4%, and 73.6%, respectively (Table 3).
Table 3
Milky ripe seed phytohormones content of the different storage period with or without peel
|
Treatment |
Determination index |
Storage time (month) |
||||
|
0 |
1 |
2 |
3 |
4 |
||
|
With peel |
GA3 (ng/g.FW) |
8.62ea |
10.53da |
16.15ca |
25.35ba |
55.07aa |
|
Without peel |
5.99eb |
7.65db |
9.13cb |
20.17bb |
55.82aa |
|
|
With peel |
ABA (ng/g.FW) |
179.97ca |
154.57ea |
211.17aa |
174.54da |
189.59ba |
|
Without peel |
129.59ab |
130.10ab |
80.53db |
83.94cb |
110.70ba |
|
|
With peel |
GA3/ABA |
0.05da |
0.07ca |
0.08cb |
0.15bb |
0.29aa |
|
Without peel |
0.05da |
0.06db |
0.11ca |
0.24ba |
0.50aa |
|
Different letters within a row indicate significant differences between the mean (p<0.05).
Seed maturity has significant influence on germination potential, germination rate, germination index and T50(d). Uniformity was not significantly influenced by seed maturity. Germination potential, germination rate and germination index were significantly affected by GA3 processing time (Table 4).
Table 4
Analysis variance the effect of GA3 processing time on germination characteristics of seeds in different maturity
|
S.O.V |
d.f. |
Germination potential (%) |
Germination rate (%) |
Germination index |
Uniformity |
T50 (d) |
|
Replication |
2 |
0.035* |
0.046** |
133.03* |
0.001 |
0.021 |
|
Seed maturity (A) |
2 |
0.573** |
0.569** |
3296.65** |
0.002 |
1.622** |
|
GA3 processing time (B) |
4 |
0.041** |
0.039** |
166.62** |
0.008ns |
0.222ns |
|
A×B |
8 |
0.31** |
0.028** |
94.07* |
0.004 |
0.122ns |
|
Error |
28 |
0.008 |
0.007 |
29.173 |
0.005 |
0.094 |
ns: non significant; *significant at 0.05 significance in F-tests; **significant at 0.001 significance in F-tests
The highest germination potential was related to full ripe seed and the lowest one was obtained by milky ripe seed; there was no significant difference between wax and full ripe seed, but both of them have significant differences with milky ripe seed. The maximum germination rate and germination index also achieved in full ripe seed. No significant differences were found in these two experimental traits between milky and ripe seed, but both of them had significant differences with full ripe seed. There were no significant differences among milky ripe seed, wax ripe seed and full ripe seed. Wax ripe seed has obtained the highest T50, but its difference with full ripe seed was not significant. However, not only wax ripe seed, but also full ripe seed had significant difference with milky seed maturity.
The maximum germination potential and germination rate was occurred in 120 and 60 min GA3 processing time, respectively. 120 min GA3 processing time had obtained the maximum germination index, which just had significant differences with control treatment (0 min). There were no significant differences among treatments in uniformity index; furthermore, the maximum one was obtained by control treatment (0 min). On the one hand, the highest T50 was related to control treatment; on the other hand, the lowest one was obtained by 60, 120 and 240 min. Moreover, there were no significant differences among treatments. The results show that seeds immersed for 60 m by GA3 had best effect to promote germination to ripe seeds, and they inhibited germination when immersed for 240 min. The maximum germination potential and germination rate was related to interaction between full ripe seed and 30 min of GA3 processing time, and the highest germination index was achieved in full ripe seed and 120 min GA3 processing time interaction. There were no significant differences among interaction traits in uniformity. Both, interaction between milky ripe seed and control treatment in processing time, interaction between milky ripe seed and 30 min of GA3 processing time had obtained the highest T50, which had significant differences with all other interaction (Table 5).
Table 5
Mean comparison for germination characteristics
|
Treatment |
Germination potential (%) |
Germination rate (%) |
Germination index |
Uniformity |
T50 (d) |
|
Seed maturity (S) |
|
|
|
|
|
|
Milky ripe seed (S1) |
38.22b |
35.47b |
21.37b |
0.9647a |
3.60a |
|
Wax ripe seed (S2) |
49.78a |
34.67b |
21.72b |
0.9520a |
3.06b |
|
Full ripe seed (S3) |
53.33a |
68.80a |
47.22a |
0.9767a |
3.00b |
|
GA3 processing time (min) (T) |
|
|
|
|
|
|
0 (T1) |
38c |
45c |
23.62b |
1.00a |
3.33ab |
|
30 (T2) |
49ab |
52c |
29.65a |
0.95a |
3.44a |
|
60 (T3) |
53a |
53a |
33.58a |
0.99a |
3.11b |
|
120 (T4) |
54a |
43d |
34.45a |
0.94a |
3.11b |
|
240 (T5) |
44bc |
28b |
29.20a |
0.93a |
3.11b |
|
A×B (S×T) |
|
|
|
|
|
|
S1T1 |
24h |
22h |
12.53e |
1.00a |
4.00a |
|
S1T2 |
29h |
24gh |
15.27de |
1.00a |
4.00a |
|
S1T3 |
49defg |
46def |
28.31c |
0.97a |
3.33b |
|
S1T4 |
52def |
50cde |
29.25c |
0.96a |
3.33b |
|
S1T5 |
37fgh |
33fgh |
21.49cde |
0.89a |
3.33b |
|
S2T1 |
21h |
21h |
13.33e |
1.00a |
3.00b |
|
S2T2 |
36fgh |
34fgh |
21.17cde |
0.93a |
3.33b |
|
S2T3 |
46efg |
45def |
28.44c |
1.00a |
3.00b |
|
S2T4 |
33gh |
33fgh |
20.64cde |
0.91a |
3.00b |
|
S2T5 |
37fgh |
38efg |
25.00cd |
0.90a |
3.00b |
|
S3T1 |
69abc |
68ab |
45.00ab |
1.00a |
3.00b |
|
S3T2 |
84a |
78a |
52.52a |
0.92a |
3.00b |
|
S3T3 |
64bcd |
64abc |
44.00ab |
1.00a |
3.00b |
|
S3T4 |
77ab |
76a |
53.46a |
0.96a |
3.00b |
|
S3T5 |
57cde |
57bcd |
41.11b |
1.00a |
3.00b |
Means with common letters within each column do not differ significantly.
Germination potential has positive and significant correlation with germination rate and germination index, which means that with increase of germination potential, germination rate increase significantly.
However, germination potential has negative and significant correlation with T50 and non-significant positive correlation with uniformity. The positive significant correlation was found between germination rate and germination index. T50 also had negative and significant correlation with both germination rate and germination index; furthermore, the correlation between uniformity and T50 was positive, but it was not significant (Table 6).
GA3 concentration had signifi-cant influence on germination potenti-al, germination rate and germination index, but uniformity and T50 were not affected by it (Table 7).
Table 6
Simple correlation among experimental characteristics in different seed maturity and GA3 processing time
|
Traits |
Germination potential |
Germination rate |
Germination index |
Uniformity |
T50 |
|
Germination potential |
1 |
|
|
|
|
|
Germination rate |
0.988** |
1 |
|
|
|
|
Germination index |
0.978** |
0.987** |
1 |
|
|
|
Uniformity |
0.009ns |
0.060ns |
0.063ns |
1 |
|
|
T50 |
-0.439** |
-0.498** |
-0.510** |
0.004ns |
1 |
ns: non significant; *significant at 0.05 significance in F-tests; **significant at 0.001 significance in F-tests
Table 7
Analysis of variance for the influence of different GA3 concentrations on wax ripe seed germination
|
S.O.V |
d.f. |
Germination potential |
Germination rate |
Germination index |
Uniformity |
T50 |
|
Replication |
2 |
0.006 |
0.006 |
16.98 |
0.012ns |
0.056 |
|
GA3 concentrations |
5 |
0.055** |
0.088** |
195.77** |
0.022ns |
0.489ns |
|
Error |
10 |
0.008 |
0.007 |
24.89 |
0.008 |
0.182 |
ns: non significant; *significant at 0.05 significance in F-tests; **significant at 0.001 significance in F-tests
The highest germination poten-tial and germination rate was related to 100 mg/l GA3 concentration, which had significant differences with all treatments, except 200 mg/l in both experimental traits.
The highest and the lowest ger-mination rate were achieved in 100 mg/l and control treatment (0 mg/l) GA3 concentration, which had significant difference with each other. 100 GA3 concentration had obtained the maximum germination index, which had significant differences with 0 mg/l and 12.5 mg/l.
There were no significant differences among treatments in uniformity. The highest T50 was related to 50 mg/l and 100 mg/l, respectively. Like uniformity, no significant difference was found among treatments (Table 8).
Table 8
Mean comparison for experimental characteristics of wax ripe seed germination in different GA3 concentration
|
Treatment |
Germination potential (%) |
Germination rate (%) |
Germination index |
Uniformity |
T50 (d) |
|
GA3 concentra-tion (mg/l) |
|
|
|
|
|
|
0 |
21.3d |
21.3c |
13.33c |
1.00a |
3.00a |
|
12.5 |
26.7cd |
26.7c |
15.99bc |
1.00a |
3.33ab |
|
25 |
37.3bcd |
49.3b |
25.75a |
0.82a |
3.33ab |
|
50 |
41.3abc |
49.3b |
25.01ab |
0.85a |
4.00a |
|
100 |
57.3a |
66.7a |
33.98a |
0.84a |
4.00a |
|
200 |
49.3ab |
53.3ab |
30.09a |
0.82a |
3.66ab |
Means with common letters within each column do not differ significantly.
Influence of different maturity seed germination by dealing with GA3
The results showed that, after manual removal of the peel, the immersed with GA3, seeds in different maturity had significantly improve-ment in germination rate, germination energy and germination index, which had significant difference with other treatments. Manual removal treatment (T1) and GA3 treatment (T2) had significant difference with CK in germination rate. Milky ripe seeds had significant differences in germination potential by dealing with GA3; seeds treated by T3 were significantly higher than other treatments in germination rate, there was no significant difference between T1 and T2, but also it was significantly higher than CK; seeds treated by T3 was significantly higher than other treatments in germination index, when others have no significant difference; T50 of T3 and T2 is shortened one day than T1 and CK. T3 had no significant difference with T2 in the germination potential of wax ripe seed, but it was significantly higher than T1 and CK; furthermore, there was no significant difference between T2 and T1, but T2 was significantly higher than CK. There was no significant difference between T1 and CK; seed treated by T3 was significantly higher than other treatments in germination rate, there was no significant difference between T1 and T2, but they were significantly higher than CK. Seed treated by T3 was significantly higher than other treatments in germination rate; furthermore, T1 was significantly higher than T2 and CK in germination index, and there was no significant difference between T2 and CK; T50 of T3 and T2 was shortened one day than T1 and CK. Full ripe seed had significant difference in germination potential. Full ripe seed had significant difference in germination rate, there was no significant difference between T3 and CK, but all of them were significantly higher than other treatments; T50 of T3 and T2 is shortened one day than T1 and CK (Table 9).
Table 9
Influence of different maturity seed germination by dealing with GA3
|
Provenances |
Treatments |
|||
|
CK |
T1 |
T2 |
T3 |
|
|
Milky ripe |
|
|
|
|
|
Germination rate (%) |
0.0d |
2.5c |
3.2b |
3.7a |
|
Germination potential (%) |
40.7c |
56.0b |
58.7b |
65.3a |
|
Germination index |
13.3b |
13.4b |
12.9b |
22.1a |
|
T50 (d) |
8 |
8 |
7 |
7 |
|
Geminating |
0.9 |
1.0 |
1.0 |
0.8 |
|
Wax ripe |
|
|
|
|
|
Germination rate (%) |
2.7c |
3.0bc |
3.5ab |
4.0a |
|
Germination potential (%) |
42.7c |
65.3b |
67.3b |
80.0a |
|
Germination index |
11.5c |
16.8b |
13.2c |
22.8a |
|
T50 (d) |
8 |
8 |
7 |
7 |
|
Geminating |
0.8 |
0.9 |
0.9 |
0.8 |
|
Full ripe |
|
|
|
|
|
Germination rate (%) |
6.7d |
8.3c |
15.3b |
22.7a |
|
Germination potential (%) |
75.7d |
77.3c |
79.3b |
92.0a |
|
Germination index |
28.1a |
20.6b |
16.1c |
29.6a |
|
T50 (d) |
7 |
7 |
6 |
6 |
Different letters within a row indicate significant differences between the mean (p<0.05)
Influence of different maturity seed germination under low temperature and room temperature storage conditions by dealing with GA3
The results showed that using GA3 treatment to harvest and storage one month cream ripe seed at room temperature, the germination rate were higher than comparison results, the difference reached significant level (p<0.05), increasing 8 and 5.1 percentage points; using GA3 treatment at room temperature to storage for 2 months, 3 months and 4 months, seed germination rate, compared to the comparison was not significant. Storage at room tempera-ture for three and four months, seed germination rate was lower than the comparison, reduced by 2 and 3 percentage points. Germination rate of the new harvest, cold storage for one and 2 months milk ripe seed treated by GA3, was higher than the comparison, the difference reached significant level (p<0.05), increasing 8, 6.9 and 5.5 percentage points, respectively; germination rate of cold storage 2 and 3 months milky ripe seed treated by GA3 was lower than the comparison for 4%, the difference was not significant (Table 10).
Table 10
Influence of milky ripe seed germination under room temperature and 0℃ storage conditions by dealing with GA3
|
Treatment |
Germination rate(%) |
||||
|
0 month |
1 month |
2 month |
3 month |
4 month |
|
|
Room temperature and no GA3 |
11e |
35.1d |
46.7c |
55b |
73a |
|
Room temperature and GA3 |
19d |
40.2c |
50.3b |
53b |
70a |
|
0℃ and no GA3 |
11e |
30.4d |
40.2c |
50b |
69a |
|
0℃ and GA3 |
19e |
37.3d |
45.7c |
51b |
65a |
Different letters within a row indicate significant differences between the mean (p<0.05)
Using GA3 treatment to newly harvested and stored one month wax ripe seeds at room temperature, the germination rate was higher than comparison results, the difference reached significant level (p<0.05), increasing by 10 and 5.5%. Using GA3 treatment at room temperature and stored for 2, 3 and 4 months, seed germination rate, relative to the comparison, was not significant, storage at room temperature for four months, seed germination rate was lower than the comparison, reducing it by 3.5%. Germination rate of the new harvest, cold storage for one and two months wax ripe seeds treated with GA3 was higher than the comparison, the difference reached significant level, increasing by 10, 7.1 and 9.6%. Germination rate of cold storage 2 and 3 months wax ripe seeds treated by GA3 was lower than the comparison by 2%, the difference was not significant (Table 11).
Table 11
Influence of full ripe seed germination under room temperature and 0℃ storage conditions by dealing with GA3
|
Treatment |
Germination rate(%) |
||||
|
0 month |
1 month |
2 month |
3 month |
4 month |
|
|
Room temperature and no GA3 |
26.3d |
50.8c |
82.2b |
92.1a |
90.3a |
|
Room temperature and GA3 |
43.2c |
78.5b |
90.5a |
90.2a |
91.6a |
|
0℃ and no GA3 |
26.3e |
45.6d |
72c |
82b |
88a |
|
0℃ and GA3 |
43.2d |
65.6c |
83b |
80ab |
85a |
Different letters within a row indicate significant differences between the mean (p<0.05).
Using GA3 treatment to harvest and storage 1 and 2 month fully mature seeds at room temperature, the germination rate was both higher than comparison results, the difference reached extremely significant level (p<0.01), increasing 16.9, 27.7 and 8.3%; using GA3 treatment at room temperature to storage for 3 and 4 months, seed germination rate compared to the comparison was not significant (p>0.05), storage at room temperature for 3 months, seed germination rate was lower than the comparison reducing it by 1.9 percentage points; germination rate of the new harvest, cold storage 1 month and 2 months fully mature seeds treated by GA3 had higher than the comparison, the difference reached significant level (p<0.05), increasing by 16.9,20 and 9%, respectively; germination rate of cold storage 3 and 4 months fully mature seeds treated by GA3 was lower than the comparison by 2% and 3%, according to storage period for the sequence, the difference was not significant (Figs. 5 and 6).
Table 12
Influence of wax ripe seed germination under room temperature and 0℃ storage conditions by dealing with GA3
|
Treatment |
Germination rate(%) |
||||
|
0 month |
1 month |
2 month |
3 month |
4 month |
|
|
Room temperature and no GA3 |
14.4e |
50.3d |
69.3c |
80b |
85.5a |
|
Room temperature and GA3 |
24.4d |
55.8c |
73.3b |
81a |
82a |
|
0℃ and no GA3 |
14.4e |
45d |
60.7c |
73b |
82a |
|
0℃ and GA3 |
24.4e |
52.1d |
70.3c |
74b |
80a |
Different letters within a row indicate significant differences between the mean (p<0.05)
The results of this experiment showed that the GA3 treatment promote seed storage at room temperature or low temperature seed germination, especially for 2 months, but not for 3 or more months or even inhibition.
GA3 and ABA interaction effects on seed germination
The test results showed the significant differences for the germination rate for each treatment, compared with CK., T5, T6, T7 (p<0.01); the T8 germination rate was significantly higher than that of T4, T3, T2, T1 (p<0.05). T1 and T2 were the most important treatment which inhibited seed germination, compared to other treatments, seed germination in T3 and T4 were 36.0% and 24.0%, respectively; under the same concentration of ABA, the seed germination rate of ABA and GA3 interaction treatment was higher than ABA treatment, but it was still lower than CK levels. These results suggest that ABA inhibits seed germination, and inhibition increased with the increase trend of ABA concentration; furthermore, GA3 can alleviate the inhibitory effect of ABA on seed germination and alleviating margin of T1, T2 with GA3 is larger than the T3, T4 (Fig.1).
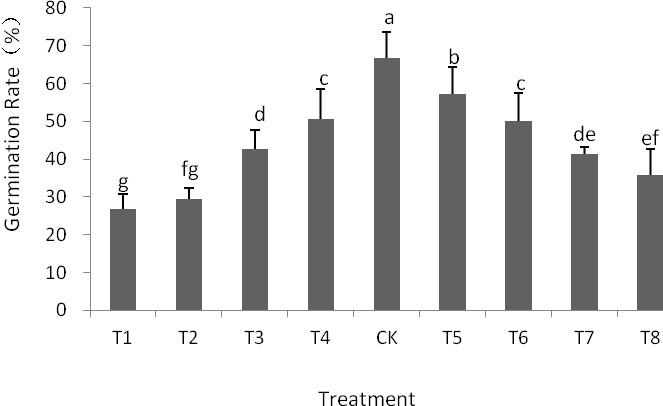
Figure 1 – GA3 and ABA interaction effects on seed germination
The results of this experiment showed that the endogenous GA3 content of each maturity degree oat seed increased with the storage period, while endogenous ABA content decreased with storage time to extend. Different maturity seed GA3/ABA ratio increase with the storage time, and this result is the same for maturity germination rate. Furthermore, GA3 content of seed without peel were higher than those with peel at each storage time, while ABA content showed the opposite. Thus, the peel may contain more ABA and conducive to seed dormancy. Lou et al. (1995) concluded that the result of the wild oat showed, small particles solid ABA/GA were higher than the large particle solid, but ABA/GA3 ratio was more greater, the lower germination rate, the higher dormancy rate. Zeng and Zhao (2001) indicated that the red string seed showed during seed development, the content of endogenous GA3 showed a decreasing trend, that GA3 content in dry seeds in room temperature and low temperature during storage have shown pre gradually reduce, the latter has an upturn, and the content of endogenous GA3 and no significant correlation with the seed germination rate. Seed germination rate significantly increased with storage time extended, the role of GA3 to enhance the seed germination rate weakened. This is similar to the results of Zeng and Zhao (2001), that related to GA3, a role on a red string seed germination. This experiment showed that when seeds are treated with different concentrations of exogenous ABA, seed germination rate decreased with the increase of ABA concentration, even when at the same time there is application of GA3, the germination rate of seed was certain upward, but still lower than in CK.
So, ABA inhibited seed germination, but its inhibitory effect in a certain extent can be remission by GA3. Wang et al. (2004) also reported the influence of ABA on the inhibition of rice seed germination results.
CONCLUSION
Seeds without peel had lower endogenous ABA content and the ratio of ABA/GA3 than seeds with peel. The best GA3 treatment time of milky ripe, wax ripe, full ripe seed were 60 min or 120 min, 60 min, and 30 min, respectively.
Seed germination rate, germination potential and germi-nation index increased before they de-clined with the increase of GA3 concentrations.
The best treatment concentration of GA3 was 100 mg/l, the turning point was 200 mg/l. The dormancy rate of low temperature storage seeds were higher than those of room temperature storage seeds at each storage time, and both decreased with the storage time.
For the seeds that were new or stored for 1-2 months, the germination rate was enhanced significantly by exogenous GA3. For the seeds that had been stored for over three months, GA3 treatment had no effect. ABA can inhibit the germination rate, which decreases with the increasing concentration of ABA.
The most inhibitive effect, which leaded to a seed germination reduction by 37.7% and 4.0%, appeared when the concentration of ABA was 500 and 1000 mg/L. GA3 could abate the effect which ABA inhibited seed germination.
Acknowledgment. We are grateful to the study grants from the earmarked fund for China Agriculture Research System (CARS-08-B-1). Thanks to the guidance of LaichunGuo and Chunlong Wang in the experiment, and thanks the workers in Baicheng Academy of Agricultural Sciences for their skilled technical assistant.
REFERENCES
Beck, E. & Ziegler, P. (1989). Biosynthesis and degradation of starch in higher plants. Ann.Rev.Plant., Physiol., 40: 95-117.
Chang, X.L., Cui, B.L., Chen, Y.L. & Xue, L.R. (2005b). Analysis the ecological environment characteristics in a farming-pasture in the Horqin Sand land. Arid Land Geograph., 28(3): 336-374 (in Chinese with English summary).
Chen, R.Z. & Huang, S.Z. (1998). Plant physiology. Zhongshan Univ. Press (in Chinese).
Chen, F., Xie, J., Zheng, W., Liu, Y., Lu, T.P., Zhao, Q., Hu, Y. & Shahrajabian, M.H. (2013). The status quo of desertification and the prevention strategy in Xinjiang. J.Food Agric.Environ., 11(2): 1025-1032.
Derera, N.F. (Ed.) (1989). The effect of pre-harvest rain. In: Pre-harvest field sprouting in cereals, CRC Press, Florida, pp. 1-14.
Facteau, T.J., Chestnut, N.E., Rowe, K.E. & Payne, C. (1992). Brine quality of gibberellic acid-treated ‘Napoleon’ sweet cherries. HortSci., 27(2): 118-192.
Hajizadeh, R. & Mirshekari, B. (2011). Interference of wild oat (Avena fatua) with wheat cultivars. JFAE, 9(3&4): 398-399.
Islam, M.R., Alam, A.M.S., Eneji, A.E., Ren, C.Z., Song, W.J. & Hu, Y.G. (2011). Evaluation of water-saving superabsorbent polymer for forage oat (Avena sativa L.) production in arid regions of northern China. J.Food Agric.Environ., 9(2): 514-518.
Khan, A.A. (1975). Primary, preventive and permissive roles of hormones in plant systems. Bot.Rev., 41(4): 391-420.
Li, M., Hu, Y.G., Zeng, Z.H., Ren, C.Z., Mao, N., Song, W.J. & Jia, P.F. (2009). Wind tunnel experiment on anti-wind erosion capacity of four crop stubbles in Horqin Sandy Land. Chinese Agric.Sci.Bull., 11: 254-258 (in Chinese with English summary).
Lin, Y.C., Zeng, Z.H., Ren, C.Z., Guo, L.C., Wang, C.L., Hlatshwayo, P. & Hu, Y.G. (2012). Effects of different soil moisture regimes on gas exchange in response to light, growth and biomass in naked oat (Avena nuda L.). J.Food Agric.Environ., 10(3&4): 1473-1478.
Lou, Y.L. & Feng, Z.H. (1995). Lixia river region of Jiangsu province of wheat following rice field oats the summering law research. Jiangsu J.Agric.Sci., 11: 25-29 (in Chinese).
Mukhtar, F.B. & Singh, B.B. (2006). Influence of photoperiod and gibberellic acid (GA3) on the growth and flowering of cowpea (Vigna unguiculata (L.) Walp). J.Food Agric.Environ., 4(2): 201-203.
Ogbaji, P.O., Shahrajabian, M.H. & Xue, X. (2013). Changes in germination and primarily growth of three cultivars of tomato under diatomite and soil materials in auto-irrigation system. Int.J.Biol., 5(3): 80, DOI: 10.5539/ijb.v5n3p80
Özkaya, O., Dündar, O. & Küden, A. (2006). Effect of preharvest gibberellic acid treatments on postharvest quality of sweet cherry. J.Food Agric.Environ., 4(1): 189-191.
Peiretti, P.G., Miraglia, N. & Bergero, D. (2011). Effects of oat or corn on the horse rations digestibility. J.Food Agric.Environ., 9(2): 268-270.
Rabiei, E., Khodambashi, M. & Pirbalouti, G. (2012). The study of the drought tolerance indices of oat (Avena sativa L.). J.Food Agric.Environ., 10(2): 646-648.
Ren, C.Z., Ma, B.L., Burrows, V., Zhou, J., Hu, Y.G., Guo, L., Wei, L., Sha, L. & Deng, L. (2007). Evaluation of early mature naked oat varieties as a summer-seeded crop in dryland northern climate regions. Field Crops Res., 103(3): 248-254, DOI: 10.1016/j.fcr.2007.07.001
Shahrajabian, M.H., Soleymani, A., Ogbaji, P.O. & Xue, X. (2017). Impact of different irrigation managements on soil water consumption, grain yield, seed protein, phosphorus and potassium of winter wheat. Cecet.Agron. in Moldova, 50(3): 5-13, DOI: 10.1515/cerce-2017-0021
Soleymani, A. & Shahrajabian, M.H. (2012). Study of cold stress on the germination and seedling stage and determination of recovery in rice varieties. Int.J.Biol., 4(4): 23, DOI: 10.5539/ijb.v4n4p23
Soleymani, A., Khoshkharam, M. & Shahrajabian, M.H. (2012). Germination rate and initial growth of silage corn grown under various fertility systems. Res. Crops, 13(3): 1035-1038.
Wang, X., Tao, L.X. & Huang, X.L. (2004). Physiological mechanisms of exogenous ABA inhibition of germination of rice seeds. Acta Agron. Sinica, 30; 1250-1253 (in Chinese).
Wilkinson, S. & Davies, W.J. (2010). Drought, ozone, ABA and ethylene: new insights from cell to plant to community. Plant Cell Environ., 33: 510-525, DOI: 10.1111/j.1365-3040.2009.02052.x
Yong, Y., Hu, Y., Shahrajabian, M.H., Ren, C., Guo, L., Wang, C. & Zeng, Z. (2017). Organic matter, protein percentage, yield, competition and economics of oat-soybean and oat-groundnut intercropping systems in Northern China. Cercet.Agron. in Moldova, 50(3): 25-35, DOI: 10.1515/cerce-2017-0023
Zeng, L. & Zhao, L.J. (2001). Effects of GA3 and ABA on seed development and germination of Salvia splendens. J. Shanghai Jiaotong Univ. (Agric.Sci.), 4: 276-279 (in Chinese).
Zhao, S.F. & Tian, C.Y. (2007). China’s oat production and research status and future direction. Rain Fed Crops, 27: 428-431 (in Chinese).

© by the authors; licensee Journal of Applied Life Sciences and Environment, Iasi, Romania. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0)
